Models
On this page you can find a description and source code of
computational models of reading and word recognition, the hippocampus,
the broader medial temporal lobe and memory systems, the eye
movement system, and of strategy analysis
for the Weather task (models are, in a sense, data analysis tools
too).
The OB1 model of reading and word recognition
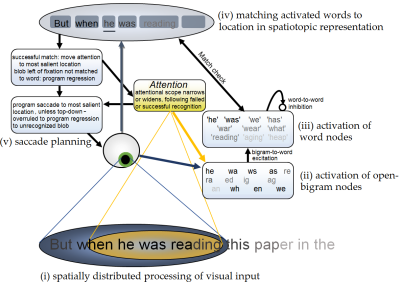
The study of reading suffers a bit from Balkanization, with every step from letter recognition to text comprehension studied in a different subfield. To reverse
this Snell, van Leipsig, Grainger and Meeter (2018)
presented OB1 Reader, the first compuational model of reading that integrates the first step
in reading (recognizing single words) with the second step, moving one's eyes through text and building up sentence representations.
Planned extensions are to address reading aloud, to understand how children learn to read and build up word representations, and have the model develop a representation of text meaning.
Summary Model of Motivation
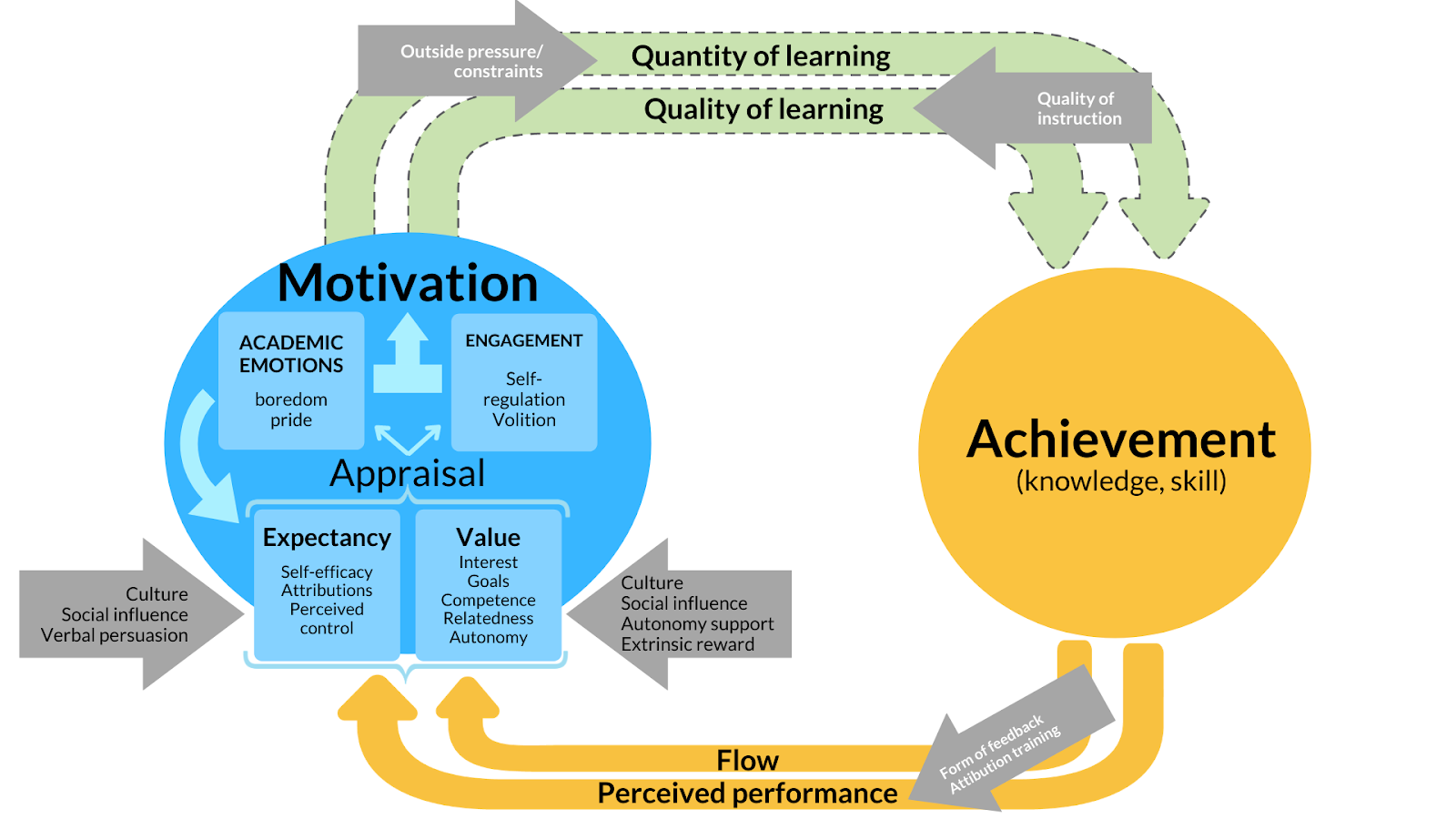
Students who are well-motivated tend to achieve more in education than students who are not.
However, that truism is only where it starts to get interesting. There is a plethora of
theories of motivation
that purport to explain how students become motivated. Although the
theories all use their own vocabulary, there are obvious similarities in how
they describe the mechanisms of motivation. With many collaborators, I therefore
made a summary model that describes how motivation and achievement interact, and what
factors support motivation. We also describe all the things that we do not yet know
about motivation-achievement interactions, which is quite a list. We elaborate on a
research agenda that may fill the blanks in our understanding of motivation and achievement.
- Blog
that explains what should count as a theory of academic motivation, and
resulting list.
- Paper
that describes summary model and research agenda.
Hippocampal physiology
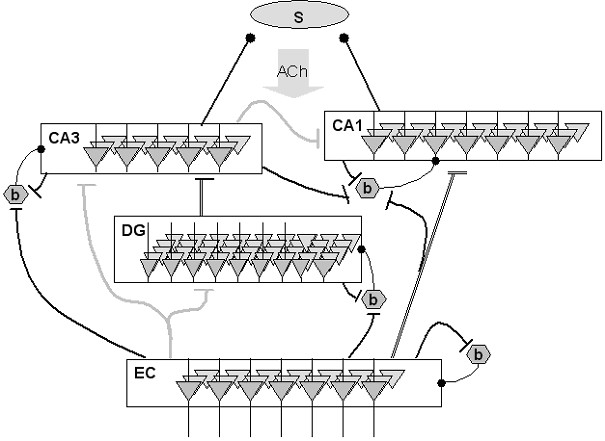
Meeter
et al. (2004) proposed a detailed model of the hippocampus
proper that focused on the dynamics of acetylcholine. It was
suggested previously by Hasselmo
and colleagues that there are two models in hippocampal
functioning: a storage mode in which information is stored without
interference from retrieved old knowledge, and a retrieval model
in which old memories are activated but new ones not laid down.
Shifting between these two modes might be under the control of
acetylcholine levels, as set by an autoregulatory
hippocampo-septo-hippocampal loop. The model investigated how such
a mechanism might operate, taking into account the major
hippocampal subdivisions, oscillatory population dynamics and the
time scale on which acetylcholine exerts its effects in the
hippocampus.
The model assumes that hippocampal mode shifting is regulated by a
novelty signal generated in the hippocampus. The simulations
suggest that this signal originates in the dentate gyrus. Novel
patterns presented to this structure lead to brief periods of
depressed firing in the hippocampal circuitry. During these
periods an inhibitory influence of the hippocampus on the septum
is lifted, leading to increased firing of cholinergic neurons. The
resulting increase in acetylcholine release in the hippocampus
causes network dynamics that favor learning over retrieval.
Resumption of activity in the hippocampus leads to the
reinstatement of inhibition.
In a later paper, Meeter
et al. (2006) adressed the effects of serotonin on memory
and the hippocampus within the same model.
Medial temporal lobe and schizophrenia
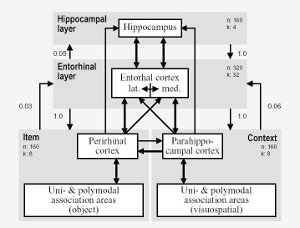
In schizophrenia, the largest cognitive impairment is usually
found in episodic memory. The cause of these deficits is unclear.
They do not seem to be related to so-called positive symptoms
(e.g., delusions), but they also do not correlate well with
negative symptoms. Talamini
et al. (2004) proposed that episodic memory impairments in
schizophrenia originate from reduced parahippocampal
connectivity. They developed a abstract medial temporal lobe model
that simulates normal performance on a variety of episodic memory
tasks. The effects of reducing parahippocampal connectivity in the
model (from perirhinal and parahippocampal cortex to entorhinal
cortex and from entorhinal cortex to hippocampus) were evaluated
and compared with findings in schizophrenia patients. Alternative
in silico neuropathologies, increased noise and loss of
hippocampal neurons, were also evaluated. In the model,
parahippocampal processing subserves integration of different
cortical inputs to the hippocampus and feature extraction during
recall. Reduced connectivity in this area resulted in a pattern of
deficits that closely mimicked the impairments in schizophrenia,
including a mild recognition impairment and a more severe
impairment in free recall. Furthermore, the "schizophrenic model"
was not differentially sensitive to interference, also consistent
with behavioral data. Neither increased noise levels, nor a
reduction of hippocampal nodes in the model, reproduced this
characteristic memory profile. Talamini
and Meeter (2009) followed up on the orignial paper by
showing that it can also explain deficits in context processing,
and later corroborated a prediction made by the model in patients
with first-episode schizophrenia (Talamini
et al, 2010).
The eye movement system
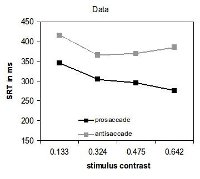

Meeter
et al. (2010) presented a model of the eye movement system in
which the programming of an eye movement is the result of the
competitive integration of information in the superior colliculi.
This brain area receives input from occipital cortex, the frontal
eye fields, and the dorsolateral prefrontal cortex, on the basis
of which it computes the location of the next saccadic target. Two
critical assumptions in the model are that cortical inputs are not
only excitatory, but can also inhibit saccades to specific
locations, and that the superior colliculi continue to influence
the trajectory of a saccade while it is being executed. With these
assumptions, we accounted for many neurophysiological and
behavioral findings from eye movement research. Interactions
within the saccade map are shown to account for effects of
distractors on saccadic reaction time and saccade trajectory,
including the global effect and oculomotor capture. In addition,
the model accounts for express saccades, the gap effect, saccadic
reaction times for antisaccades, and recorded responses from
neurons in the superior colliculi and frontal eye fields in these
tasks. Later, we showed how this model can account for saccades
that deviate away from distractors, which could until then only be
explained through implausible assumptions about inhibition (Kruijne et
al., 2014).
Entry into working memory and the attentional
blink
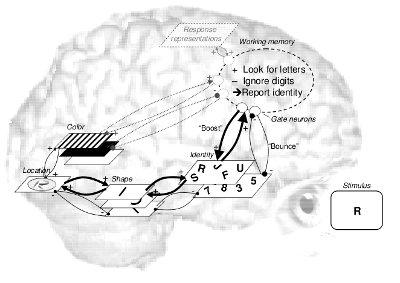
What is the time course of visual attention? Attentional blink
studies have found that the second of two targets is often missed
when presented within about 500 ms from the first target,
resulting in theories about relatively long-lasting capacity
limitations or bottlenecks. Earlier studies, however, have
reported quite the opposite finding: Attention is transiently
enhanced, rather than reduced, for several hundreds of
milliseconds after a relevant event. Olivers
and Meeter (2008) present a general theory as well as a working
computational model which integrate these findings. There is no
central role for capacity limitations or bottlenecks. Central is a
rapidly responding gating system (or attentional filter) that
seeks to enhance relevant and suppress irrelevant information.
When items sufficiently match the target description, they elicit
transient excitatory feedback activity (a "boost" function), meant
to provide access to working memory. However, in the attentional
blink task, the distractor after the target is accidentally
boosted, resulting in subsequent strong inhibitory feedback
response (a "bounce"), which in effect closes the gate to working
memory. The theory explains many findings that are problematic for
limited-capacity accounts, including a new experiment showing that
the attentional blink can be postponed.
Multiple memory stores and classical
conditioning
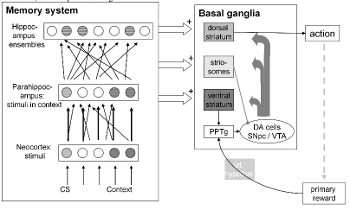
Why does the brain contain more than one memory system? What's
the relation between all these systems? Meeter
et al. (2004) do not give much of a rationale, but their
computational model of memory-oriented brain regions contains some
organizing principles. They dinstinguish between a hierarchy of
episodic memory systems, in which the main difference between
stores is the level of integration of representations: from single
stimuli in the cortex to integrated represations of the whole
situation in the hippocampus. These stores are inputs to a number
of output regions that themselves store associations between
imputs and behavioral responses, such as with fear responses in
the amygdala. In Meeter et al. (2004), findings from classical
conditioning are analyzed within the model. It describes how a
familiarity signal may arise from parahippocampal cortices, giving
a novel explanation for the finding that the neural response to a
stimulus in these regions decreases with increasing stimulus
familiarity. Recollection is ascribed to the hippocampus proper.
It is shown how the properties of episodic representations in the
neocortex, parahippocampal gyrus and hippocampus proper may
explain phenomena in classical conditioning. The model reproduces
the effects of hippocampal, septal, and broad hippocampal region
lesions on contextual modulation of classical conditioning,
blocking, learned irrelevance, and latent inhibition.
In a later paper (Meeter
et al, 2009), genetic algorithms to investigate the
complexity of memory. Model animals were constructed containing a
dorsal striatal layer that controlled actions, and a ventral
striatal layer that controlled a dopaminergic learning signal.
Both layers could gain access to the Meeter et al. (2004) memory
hierarchy, but such access was penalized as energy expenditure.
Model animals were then selected on their fitness in simulated
operant conditioning tasks. Results suggest that having access to
multiple memory stores and their representations is important in
learning to regulate dopamine release, as well as in contextual
discrimination. For simple operant conditioning, as well as
stimulus discrimination, hippocampal compound representations
turned out to suffice, a counterintuitive result given findings
that hippocampal lesions tend not to affect performance in such
tasks. However, there is in fact evidence to support a role for
compound representations and the hippocampus in even the simplest
conditioning tasks.
Strategy analysis for Weather
prediction & other multicue learning tasks
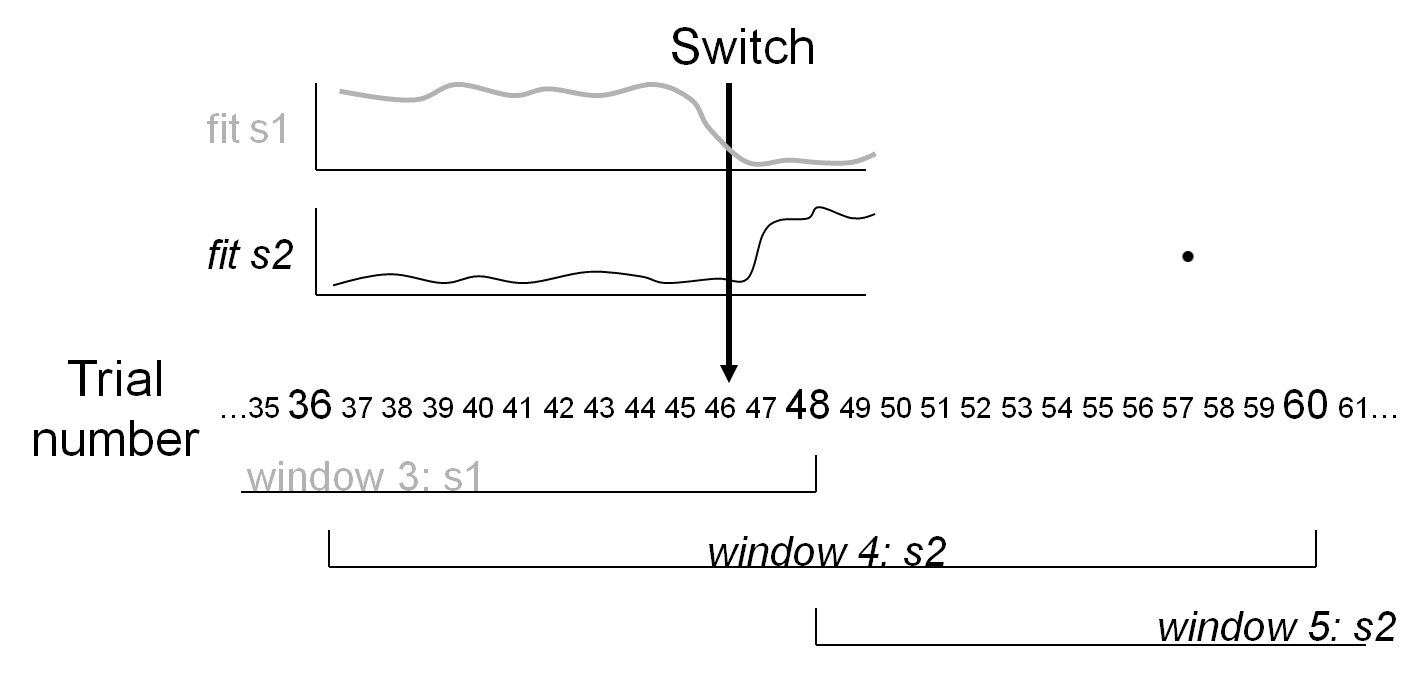 The "weather prediction" task is
a widely used task for investigating probabilistic category
learning, in which various cues are probabilistically (but not
perfectly) predictive of class membership. This means that a given
combination of cues sometimes belongs to one class and sometimes
to another. Prior studies showed that subjects can improve their
performance with training, and that there is considerable
individual variation in the strategies subjects use to approach
this task. Strategy analysis of probabilistic categorization
attempts to identify the strategy followed by a participant. Monte
Carlo simulations show that the analysis can indeed reliably
identify such a strategy if it is used, and can identify switches
from one strategy to another (Meeter
et al, 2006). Analysis of data from normal young adults
shows that the fitted strategy can predict subsequent responses.
Moreover, learning is shown to be highly nonlinear in
probabilistic categorization. Analysis of performance of patients
with dense memory impairments due to hippocampal damage shows that
although these patients can change strategies, they are as likely
to fall back to an inferior strategy as to move to more optimal
ones (Meeter
et al, 2008).
The "weather prediction" task is
a widely used task for investigating probabilistic category
learning, in which various cues are probabilistically (but not
perfectly) predictive of class membership. This means that a given
combination of cues sometimes belongs to one class and sometimes
to another. Prior studies showed that subjects can improve their
performance with training, and that there is considerable
individual variation in the strategies subjects use to approach
this task. Strategy analysis of probabilistic categorization
attempts to identify the strategy followed by a participant. Monte
Carlo simulations show that the analysis can indeed reliably
identify such a strategy if it is used, and can identify switches
from one strategy to another (Meeter
et al, 2006). Analysis of data from normal young adults
shows that the fitted strategy can predict subsequent responses.
Moreover, learning is shown to be highly nonlinear in
probabilistic categorization. Analysis of performance of patients
with dense memory impairments due to hippocampal damage shows that
although these patients can change strategies, they are as likely
to fall back to an inferior strategy as to move to more optimal
ones (Meeter
et al, 2008).
Strategy analysis can be done in excel with the file below.
Matlab code is also available upon request.